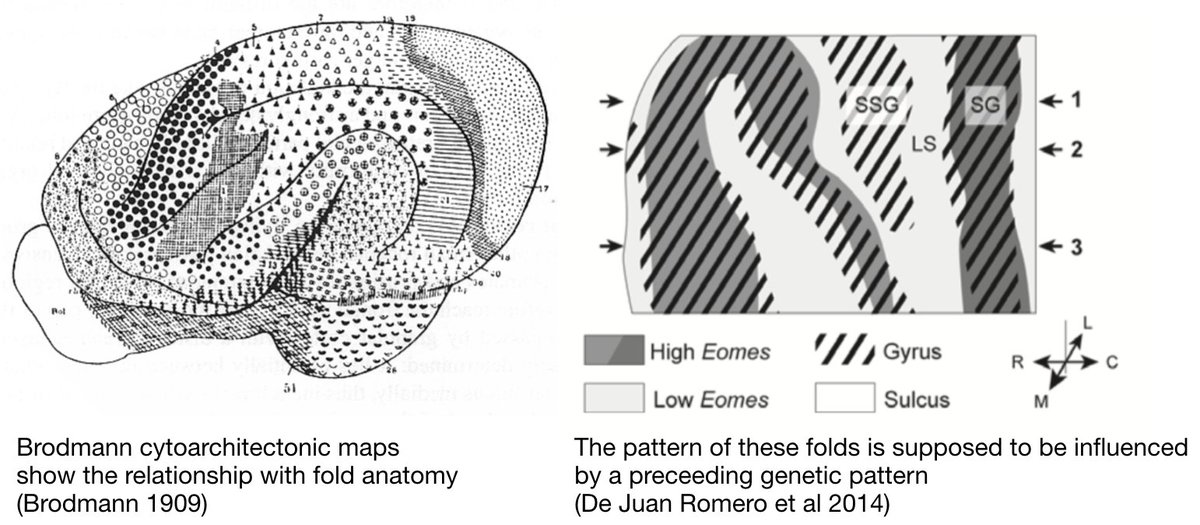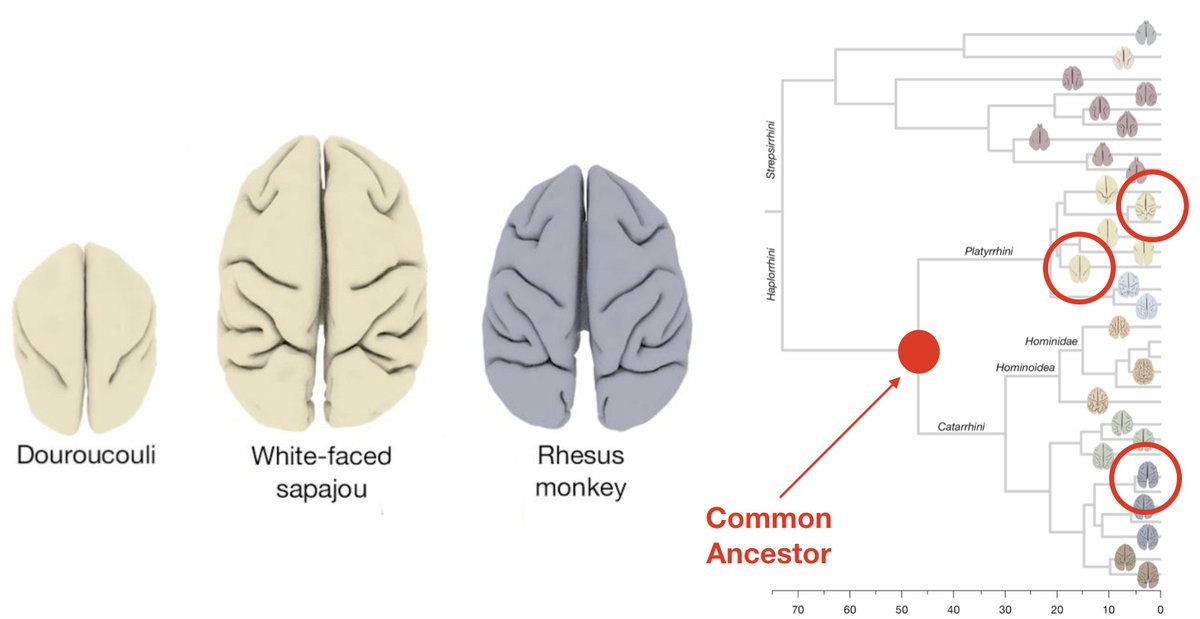
1. The particular type of causal regime at work in biological organisation is 𝘾𝙡𝙤𝙨𝙪𝙧𝙚. It combines:
* An open thermodynamic regime, and
* A closed organisation regime of 𝙘𝙤𝙣𝙨𝙩𝙧𝙖𝙞𝙣𝙩𝙨
* An open thermodynamic regime, and
* A closed organisation regime of 𝙘𝙤𝙣𝙨𝙩𝙧𝙖𝙞𝙣𝙩𝙨

2. 𝘽𝙞𝙤𝙡𝙤𝙜𝙞𝙘𝙖𝙡 𝙙𝙚𝙩𝙚𝙧𝙢𝙞𝙣𝙖𝙩𝙞𝙤𝙣 𝙖𝙨 𝙨𝙚𝙡𝙛-𝙘𝙤𝙣𝙨𝙩𝙧𝙖𝙞𝙣𝙩
Closure can be defined in different contexts:
i. In general, a domain K is said to have closure if all operations defined in it remain within the same domain.
Closure can be defined in different contexts:
i. In general, a domain K is said to have closure if all operations defined in it remain within the same domain.
ii. For a system, its operation is said to realise closure if the results of its action remain within the system (Bourgine and Varela 1992).
mitpress.mit.edu/books/toward-p…
mitpress.mit.edu/books/toward-p…
iii. For an organism, closure is realised if:
1. Processes are related as a network, so that they recursively depend on each other in their own realisation, and
2. They constitute a system as a unity recognisable in space in which the processes exist
doi.org/10.1002/bs.383…
1. Processes are related as a network, so that they recursively depend on each other in their own realisation, and
2. They constitute a system as a unity recognisable in space in which the processes exist
doi.org/10.1002/bs.383…
5. Whereas autopoiesis is defined as closure of processes, here it's defined at the level of constraints: local and contingent causes, exerted by specific structures/processes, reducing the degrees of freedom of the system on which they act doi.org/10.1007/978-94…
@carl_b_sachs
@carl_b_sachs
6. Autonomous systems are capable of self-constraint: closed at the level of constraints, but thermodynamically open.
7. 𝙏𝙝𝙚 𝙩𝙝𝙚𝙧𝙢𝙤𝙙𝙮𝙣𝙖𝙢𝙞𝙘 𝙜𝙧𝙤𝙪𝙣𝙙𝙞𝙣𝙜 𝙤𝙛 𝙖𝙪𝙩𝙤𝙣𝙤𝙢𝙮
Biological organisation has to be understood from its thermodynamic grounding – a balance between
i) components used to produce energy, and
Biological organisation has to be understood from its thermodynamic grounding – a balance between
i) components used to produce energy, and
ii) those able to modify rates of reaction ensuring synchronisation of couplings
8. In a biol. system constraints are required to harness the flow of energy (the system doesn’t only generate heat). They are also 𝙘𝙤𝙣𝙨𝙩𝙞𝙩𝙪𝙩𝙞𝙫𝙚 when they're at the same time the condition and the product of the system’s work (Kauffman 2000) global.oup.com/academic/produ…
9. Kauffman's approach misses, however, one aspect: organisational closure requires not only the constraining action exerted on the thermodynamic flow, but also a 𝙨𝙥𝙚𝙘𝙞𝙛𝙞𝙘 𝙤𝙧𝙜𝙖𝙣𝙞𝙨𝙖𝙩𝙞𝙤𝙣 among the constitutive constraints (namely, closure of constraints).
10. 𝘾𝙤𝙣𝙨𝙩𝙧𝙖𝙞𝙣𝙩𝙨 𝙖𝙣𝙙 𝙥𝙧𝙤𝙘𝙚𝙨𝙨𝙚𝙨
𝙋𝙧𝙤𝙘𝙚𝙨𝙨𝙚𝙨 are the whole set of physicochemical changes
𝘾𝙤𝙣𝙨𝙩𝙧𝙖𝙞𝙣𝙩𝙨 are entities which act on these processes while remaining unaffected by them and the processes’ time scale.
#definition
𝙋𝙧𝙤𝙘𝙚𝙨𝙨𝙚𝙨 are the whole set of physicochemical changes
𝘾𝙤𝙣𝙨𝙩𝙧𝙖𝙞𝙣𝙩𝙨 are entities which act on these processes while remaining unaffected by them and the processes’ time scale.
#definition
Constraints can be defined in terms of symmetry:
C is a constraint on a process A➾B (produce B from A) at a timescale 𝜏 if:
i) A➾B and Ac➾Bc are not symmetrical. In other words, the result of converting A into B is affected by the presence of the constraint C
C is a constraint on a process A➾B (produce B from A) at a timescale 𝜏 if:
i) A➾B and Ac➾Bc are not symmetrical. In other words, the result of converting A into B is affected by the presence of the constraint C
ii) There's a temporal symmetry associated with C_{A➾B} in relation to the process Ac=>>Bc at time scale 𝜏. In other words, the occurrence of the obtention of B from A does not affect C: C is conserved.
11. In most biol. systems, constraints alter their behaviour but don't lead to new behaviours:
B could be produced by A without C, although that may be very unlikely.
Constraints do not extend the space of possible dynamics, just shape their probability!
B could be produced by A without C, although that may be very unlikely.
Constraints do not extend the space of possible dynamics, just shape their probability!
https://twitter.com/Colinoscopy/status/1255890780641689601?s=20&t=84yrmCnBVYAuWcKuEVOMuQ
12. Constraints may be 𝙚𝙣𝙖𝙗𝙡𝙞𝙣𝙜 or 𝙡𝙞𝙢𝙞𝙩𝙞𝙣𝙜. F. ex, enabling at short time scale (contribute to make happen something that would be otherwise almost impossible), and limiting (canalising) at long time scales.
mitpress.mit.edu/books/dynamics…
mitpress.mit.edu/books/dynamics…
13. The symmetry condition (ii) implies that no relevant flow of matter or energy occurs between C_{A➾B} and A➾B, hence, at time scale 𝜏, constraints can be treated as if they were not thermodynamic objects!
14. The central difference between constraints and processes is that they correspond to two different causal regimes:
- Processes are a 𝙢𝙖𝙩𝙚𝙧𝙞𝙖𝙡 cause,
- Constraints are an 𝙚𝙛𝙛𝙞𝙘𝙞𝙚𝙣𝙩 cause (at timescale 𝜏).
plato.stanford.edu/entries/aristo…
- Processes are a 𝙢𝙖𝙩𝙚𝙧𝙞𝙖𝙡 cause,
- Constraints are an 𝙚𝙛𝙛𝙞𝙘𝙞𝙚𝙣𝙩 cause (at timescale 𝜏).
plato.stanford.edu/entries/aristo…
15. 𝙁𝙧𝙤𝙢 𝙨𝙚𝙡𝙛-𝙤𝙧𝙜𝙖𝙣𝙞𝙨𝙖𝙩𝙞𝙤𝙣 𝙩𝙤 𝙗𝙞𝙤𝙡𝙤𝙜𝙞𝙘𝙖𝙡 𝙤𝙧𝙜𝙖𝙣𝙞𝙨𝙖𝙩𝙞𝙤𝙣
* Self-determination: broader idea of closed causal loop
* Self-organisation: type of self-determination found in the physical and chemical domain
#definition
* Self-determination: broader idea of closed causal loop
* Self-organisation: type of self-determination found in the physical and chemical domain
#definition
* Dissipative structures: example of self-organisation, such as Bénard cells, flames, hurricanes, oscillatory chemical reactions, ...
#definition
youtube.com/clip/UgkxODG6s…
#definition
youtube.com/clip/UgkxODG6s…
16. Dissipative systems realise a minimal form of self-determination. They generate a single macroscopic structure, unlike biol. systems which can exert a high number of constraints & generate networks of structures realising collective self-maintenance.
17. The distinction between self-organisation and closure involves the takeover of the boundary conditions required for the maintenance of the system.
Self-organising systems are not just too simple: they are not a relevant starting point for the emergence of biological autonomy.
Self-organising systems are not just too simple: they are not a relevant starting point for the emergence of biological autonomy.
18. 𝘿𝙚𝙥𝙚𝙣𝙙𝙚𝙣𝙘𝙚
A single constraint C acting on the process A➾B at timescale 𝜏 can be written C(A➾B)𝜏 and represented diagrammatically as
A single constraint C acting on the process A➾B at timescale 𝜏 can be written C(A➾B)𝜏 and represented diagrammatically as

19. There is dependence among constraints when the replacement or repair of a constraint depends on the action of another: C1 is a constraint at scale 𝜏1, C2 is a constraint at scale 𝜏2 or a process producing aspects of C1 relevant for its role as constraint as scale 𝜏1

20. 𝘾𝙡𝙤𝙨𝙪𝙧𝙚
Is the mutual dependence of constraints:
Is the mutual dependence of constraints:

*Closure is not independence
*Closure is a general invariant of biological organisation
*Closure is a fundamental principle of order of biological phenomena:
*Closure is a general invariant of biological organisation
*Closure is a fundamental principle of order of biological phenomena:

21. 𝘼 𝙬𝙤𝙧𝙙 𝙖𝙗𝙤𝙪𝙩 𝙧𝙚𝙡𝙖𝙩𝙚𝙙 𝙢𝙤𝙙𝙚𝙡𝙨
Time to acknowledge the pioneering work of Maturana & Varela’s autopoiesis, Pattee’s concept of constraint, Szathmary’s reflexive catalysis, Kauffman’s catalytic closure, Gánti’s chemoton, Rosen’s relational biology 🙏
Time to acknowledge the pioneering work of Maturana & Varela’s autopoiesis, Pattee’s concept of constraint, Szathmary’s reflexive catalysis, Kauffman’s catalytic closure, Gánti’s chemoton, Rosen’s relational biology 🙏
22. 𝙍𝙚𝙜𝙪𝙡𝙖𝙩𝙞𝙤𝙣
Regulation addresses variations which call for a response by the global organisation, unlike Constitutive stability, which is related to local robustness (f.ex., in systems with negative feedback loops such as Gánti’s chemoton).
Regulation addresses variations which call for a response by the global organisation, unlike Constitutive stability, which is related to local robustness (f.ex., in systems with negative feedback loops such as Gánti’s chemoton).
23. Constitutive stability is an example of 𝙧𝙤𝙗𝙪𝙨𝙩𝙣𝙚𝙨𝙨, and alone does not enable the system to explore different regimes of closure.
On the contrary, Regulatory constraints:
On the contrary, Regulatory constraints:
*Don’t contribute to the maintenance of closure in stable conditions, but when the system is disrupted, they govern its transition towards re-establishment.
*Are not subject to constitutive closure: they are 2nd-order in the sense that they act on other constitutive constraints
*Are not subject to constitutive closure: they are 2nd-order in the sense that they act on other constitutive constraints
*Realise a sort of decoupling from the constitutive organisation
*Produce reversible actions: the system can be transitioned back to its original state
*Produce reversible actions: the system can be transitioned back to its original state
24. Regulation is not only robust (like constitutive stability) but adaptive; regulates 1st-order norms according to 2nd-order norms; is what characterises metabolism (and not only constitutive stability)
25. Regulation is what allows stability while enabling an increase in complexity, by enabling the composition of constitutive building blocks.
26. 𝙏𝙤𝙬𝙖𝙧𝙙𝙨 𝙖𝙪𝙩𝙤𝙣𝙤𝙢𝙮
Regulation represents a qualitative transition on the path towards autonomy. With regulated closure, a system can generate intrinsic norms according to 2n-order norms.
However, we still need an 𝙞𝙣𝙩𝙚𝙧𝙖𝙘𝙩𝙞𝙫𝙚 𝙙𝙞𝙢𝙚𝙣𝙨𝙞𝙤𝙣.
Regulation represents a qualitative transition on the path towards autonomy. With regulated closure, a system can generate intrinsic norms according to 2n-order norms.
However, we still need an 𝙞𝙣𝙩𝙚𝙧𝙖𝙘𝙩𝙞𝙫𝙚 𝙙𝙞𝙢𝙚𝙣𝙨𝙞𝙤𝙣.
27. End of Chapter 1.
Continue to Chapter 2!
Continue to Chapter 2!
https://twitter.com/R3RT0/status/1525467536678084608?s=20&t=wrI1bJ3nXcBvje_ay_PSCg
• • •
Missing some Tweet in this thread? You can try to
force a refresh