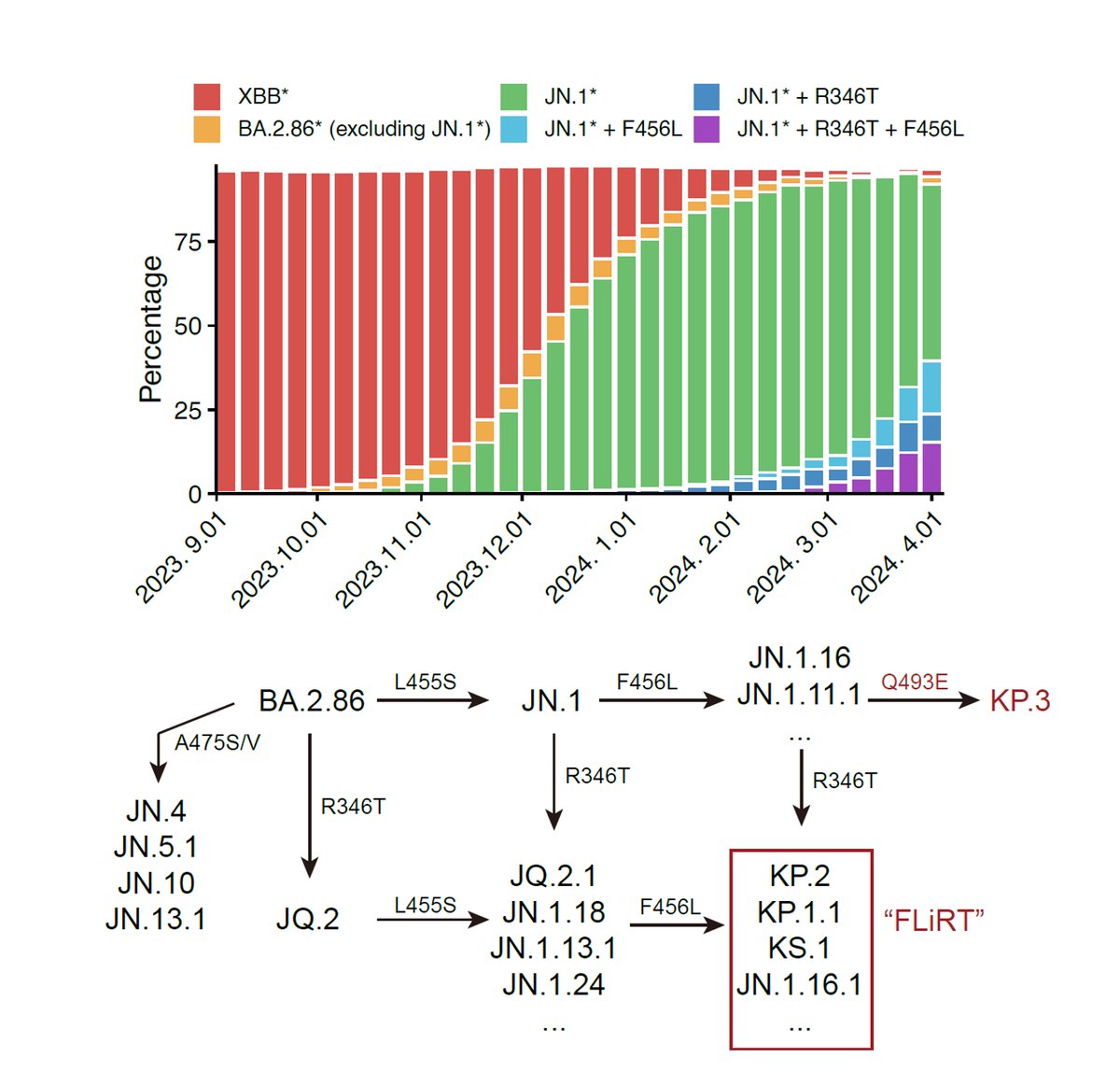
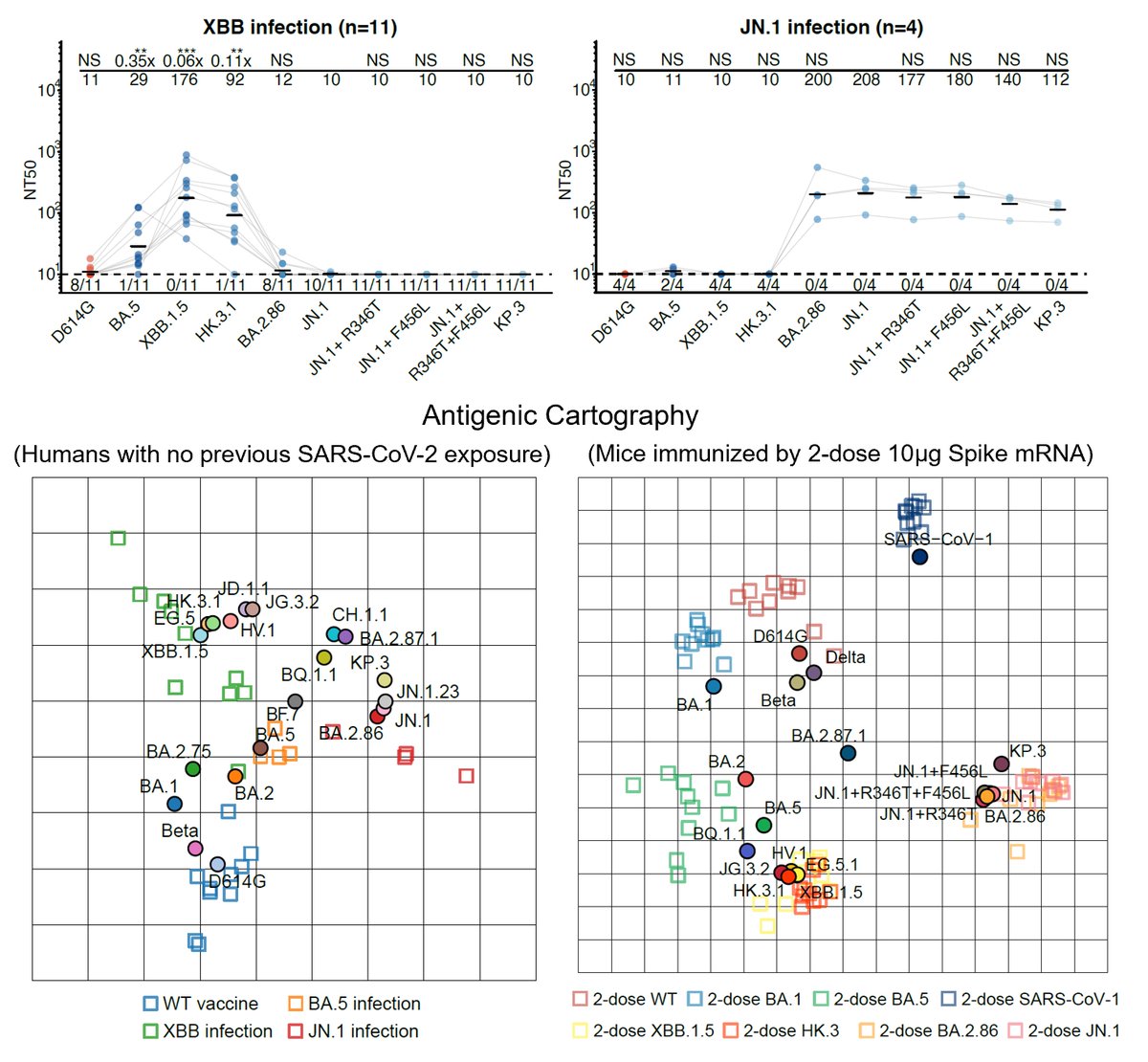
Our latest preprint is out, where we investigated a profound SARS-CoV-2 epidemiological anomaly: BA.3.2.2 is selectively infecting children. Here, we show that the lack of ancestral-strain immune imprinting is promoting BA.3.2.2 pediatric infections.
1/9
biorxiv.org/content/10.648…
1/9
biorxiv.org/content/10.648…

We find that children under 8 with no history of SARS-CoV-2 vaccination have very limited neutralization against BA.3.2.2. This is in stark contrast to children who have received the ancestral-strain vaccines (imprinted). These findings also echo David Ho's data.
2/9
2/9

We found that this imprinting dependency also holds true in adults. Weakly imprinted inactivated vaccine recipients exhibit significantly lower BA.3.2.2 neutralization titers than strongly imprinted mRNA-vaccinated individuals.
3/9
3/9

To validate this on a larger scale, we stratified a large inactivated-vaccinated cohort into highly imprinted vs weakly imprinted groups based on their NT50s fold-changes relative to D614G. The conclusion holds: weaker imprinting leads to reduced BA.3.2.2 neutralization.
4/9
4/9

Single-B-cell V(D)J sequencing reveals the mechanistic driver: strong ancestral imprinting enriches for IGHV3-53/66 public Class 1 antibodies (efficient against BA.3.2.2), whereas weak imprinting forces reliance on IGHV2-5/5-51 Class 1/4 antibodies (escaped by BA.3.2.2).
5/9
5/9

The evolutionary risk: if BA.3.2.2 scales up, Class 1 evading mutations like F456L or the 455-456 "FLip" combo are highly tolerated and maintain viral fitness. This would allow the lineage to erode the residual neutralizing immunity of strongly imprinted adults.
6/9
6/9

This means that the sustained transmission in children may catalyze BA.3.2.2 variants that combine pediatric-evading features with adult-evading Class 1 mutations (like F456L). This could allow the lineage to breach adult imprinted immunity and drive global transmission.
7/9
7/9
Key takeaways: Classic imprinting (original antigenic sin) is usually seen as a constraint, but here it serves as a protective factor. The functional outcome of imprinting should always be defined by the specific interactions between antigens and the host B cell repertoire.
8/9
8/9
We might also need to rethink pediatric SARS-CoV-2 vaccination. Future strategies may need to weigh the immediate benefits of variant-matched antigens against the long-term necessity of establishing a broad antibody baseline to ensure balanced protection across all ages.
9/9
9/9
• • •
Missing some Tweet in this thread? You can try to
force a refresh