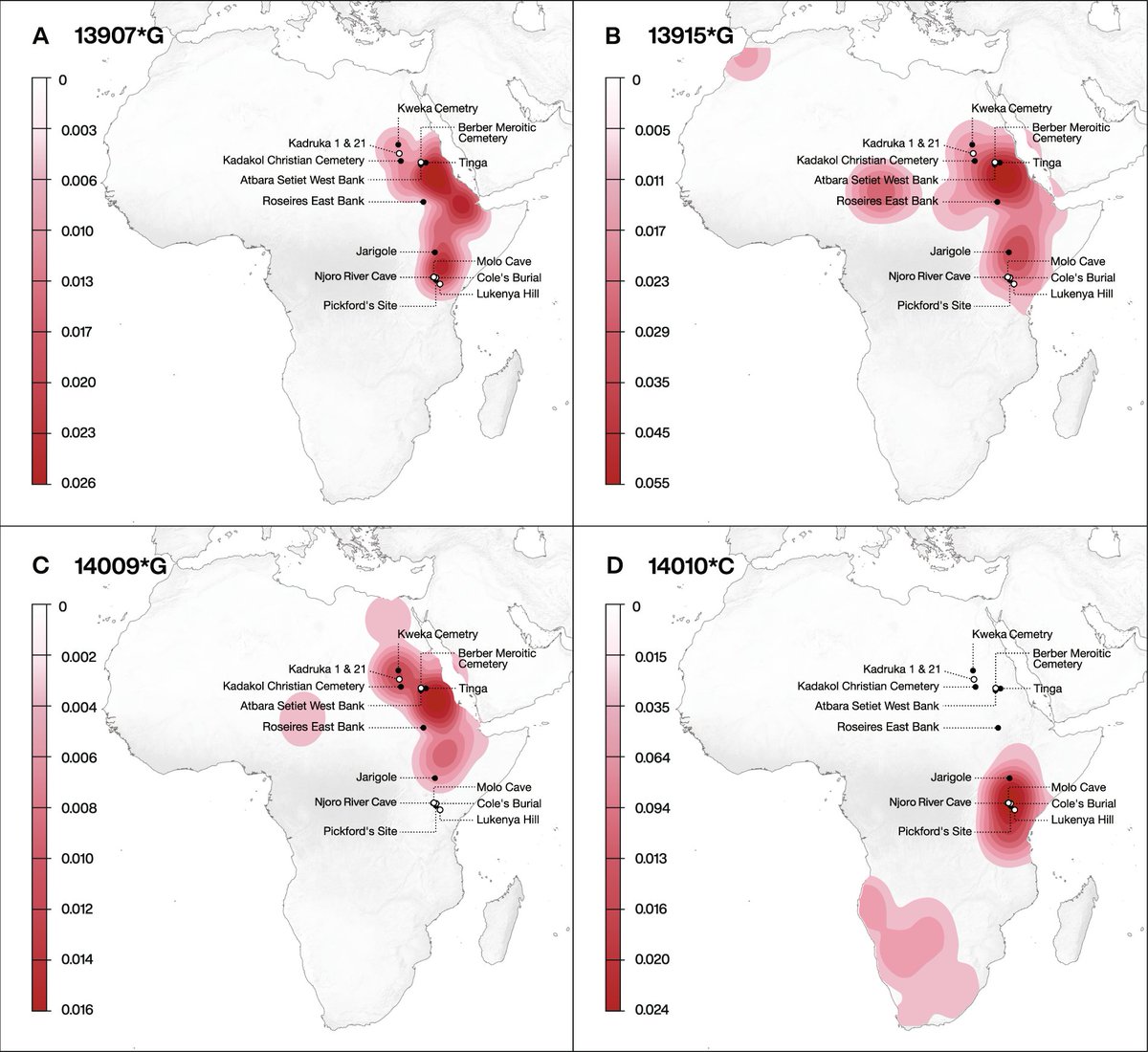
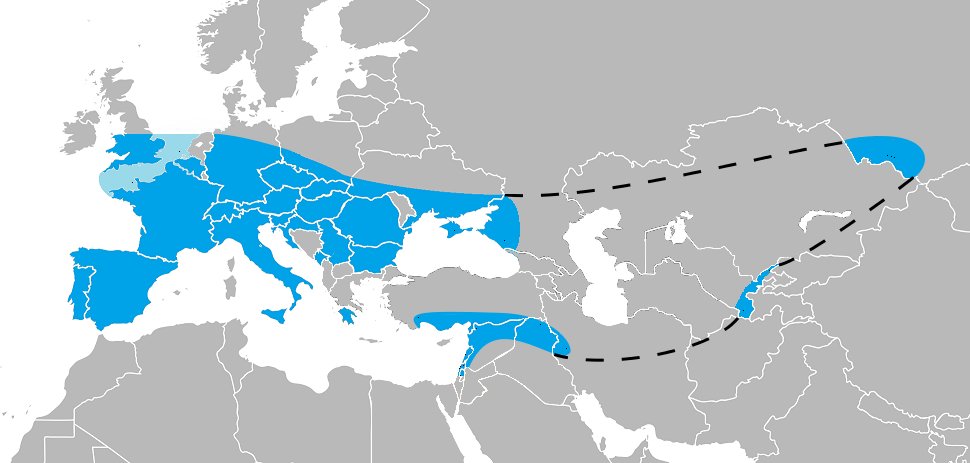
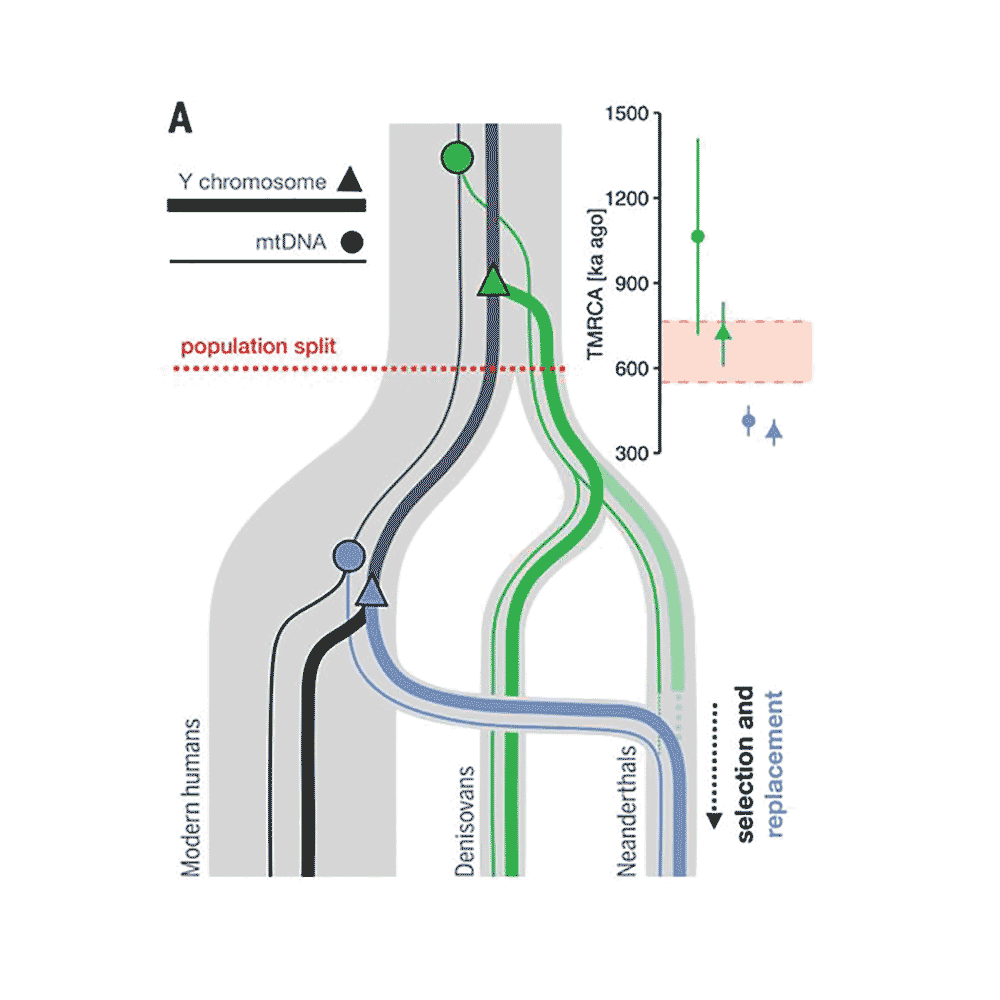
A lot of problems in science come down to whether larger amounts of noisy data are better or worse than smaller amounts of high-precision data. Of course, in paleoanthropology we usually are faced with small amounts of noisy data.
Our problems are that paleoanthropologists make up for shortfalls in data by carting in models and assumptions. These take on a life of their own, so much that even new discoveries that provide high-precision data cannot make a dent in most people's research direction.
I'm reading very carefully today an article by @Keilmesser and Marlize Lombard, which presents a very conventional view that large-brained hominins must have made MSA toolkits and smaller-brained hominins could not have done so. It's a thoughtful paper, but I'm unpersuaded.
The paper is in the Journal of Archaeological Method and Theory, here: doi.org/10.1007/s10816… I would advocate a more critical examination of noisy data. For example, I don't think the data show whether or not H. naledi and H. sapiens (or other hominins) were sympatric.
The argument that larger brain size supplies the "cognitive requirements of lithic technology" depends implicitly on the assumption that tools come from domain-general cognitive mechanisms. We increasingly see in comparative and experimental psychology that this isn't true.
Stone age archaeology is long overdue for a re-evaluation of the concept of "association". Finding a bone within a geological stratum that also contains stones links them only with the temporal resolution of the stratum -- which may extend tens to hundreds of thousands of years.
This paper grapples with the problem of niche differentiation, and like earlier work on Neandertals and modern humans, it is premised on the idea that one species is dominating a cognitive niche and the other pecking at the edges. That's not differentiation, that's domination.
Obviously I appreciate the thoughtful and deep reading of our work on H. naledi, and our engagement with these arguments must take a much longer form. For now, I caution against the "feel-good" maintenance of the "modern human revolution" viewpoint that this paper represents.
• • •
Missing some Tweet in this thread? You can try to
force a refresh